Nelson Fernandes Gomes - São Paulo, 1991
 |
| Nesta foto: Monodelphis macae Gomes, 1991 |
| CONTEÚDO | |||
Introdução
Revisão do Gênero Monodelphis
Organização da pelagem
Caracteres cranianos
Dentição
Anatomia externa (Rinário, orelha, patas e cauda.)
Medidas corporais
Estudo do crânio
Medidas variáveis com o crescimento
Medidas que não variam com o crescimento
Tabelas 1-10
Chaves de identificação com figuras (peles e crânios)
Caracteres cranianos
Dentição
Anatomia externa (Rinário, orelha, patas e cauda.)
Medidas corporais
Estudo do crânio
Medidas variáveis com o crescimento
Medidas que não variam com o crescimento
Tabelas 1-10
Chaves de identificação com figuras (peles e crânios)
Grupo Monodelphis domestica
Grupo Monodelphis brevicaudata
Grupo Monodelphis dimidiata
Grupo Monodelphis emiliae
Grupo Monodelphis americana
Grupo Monodelphis adusta
Filogenia
BiogeografiaGrupo Monodelphis brevicaudata
Grupo Monodelphis dimidiata
Grupo Monodelphis emiliae
Grupo Monodelphis americana
Grupo Monodelphis adusta
Filogenia
Conclusões
Summary
Literatura citada
INTRODUÇÃO
O gênero Monodelphis Burnett 1830 (Mammalia: Marsupialia: Didelphimorphia: Didelphidae: Monodelphinae) distinguem-se dos demais marsupiais americanos por sua cauda relativamente curta em relação ao comprimento do corpo (aproximadamente metade do comprimento da cabeça e do tronco juntos), tamanho pequeno (variando entre 10-28 cm), orelhas curtas, hálux mais curto e mais paralelo em relação aos outros artelhos, almofadas palmares e plantares proporcionalmente menores e manus modificada com garras mais alongadas. As espécies desse gênero possuem hábitos terrestres, abrigando-se em tocas e alimentando-se de insetos (Goeldi, 1894: 465), Cabrera (1940: 29), algumas espécies podem ser diurnas ou noturnas, com atividade predominantemente terrestre e crepuscular Eisenberg (1999: 67). Goeldi, (1894: 465) relatou que M. americana tem hábitos preferencialmente noturnos. A sua anatomia externa, ou seja, a cauda curta, as orelhas pequenas e a posição do hálux, sugerem uma adaptação semi-fossorial, o que vem sendo confirmado por estudos nesse sentido.
O gênero é encontrado em florestas úmidas, campos cerrados, campos abertos e em regiões semi-áridas como a Caatinga do nordeste brasileiro e o Chaco argentino-paraguaio, Myers & Wetzel (1974). Monodelphis é um gênero praticamente sulamericano, sua distribuição geográfica tem como limite o extremo leste do Panamá, estendendo-se para o sul até o norte da Argentina. O gênero é predominantemente brasileiro, uma única espécie, não tem chances de ocorrer no Brasil: Monodelphis osgoodi, que é uma espécie das altitudes andinas da Bolívia e Peru. As demais certamente ocorrem, ainda que M. reigi não tenha sido registrada.
A primeira referência na literatura a uma espécie desse gênero foi de Marcgrave (1648: 229) como Mus araneus (rato aranha), citando que assemelhava-se ao rato. A segunda referência é a de Albertus Seba (1734: 50) como Muris sylvestris Americani foemina (rato selvagem da América, fêmea), também uma referência ao rato. Buffon (1767: 160), baseado em Marcgrave (1648), registrou "La Musaraigne du Brèzil", associando o gênero a um musaranho, copiado mais tarde por Pennant (1777: 309) como "Brazilian Shrew". O primeiro nome reconhecido oficialmente para uma espécie desse gênero foi Sorex americanus Muller (1776: 36) (=Monodelphis americana) com base na descrição de Buffon (1767: 160). O segundo nome reconhecido foi Didelphis brevicaudata Erxleben (1777: 80) (=Monodelphis brevicaudata), criada com base na descrição de Seba (1734: 50).
Inicialmente, esse conjunto de espécies foi alocado no gênero Didelphis de Lineu. Posteriormente, essas espécies foram colocadas no gênero Peramys Lesson (1842: 187) e foram assim tratadas como um subgênero de Didelphis por Thomas (1888a), que forneceu uma lista de dez espécies, quase todas ainda dentro de Monodelphis. Entretanto, Burnett (1830: 351) ainda que de uma maneira leviana (com uma interrogação: Monodelphis?) já havia proposto o status genérico atual, que só mais tarde foi reconhecido.
Cabrera (1958), apresentou uma lista com dezoito formas, entre espécies e subspécies, além de listas com sinonímias e distribuições geográficas, que foram por longo tempo usadas como única referência sobre o grupo. Também muito significativo foi o trabalho de Pine, que entre 1975 e 1984, publicou notas específicas sobre problemas existentes na sistemática de Monodelphis, ajudando a esclarecer o status taxonômico de algumas espécies.
Eisenberg & Redford (1989, 1992 e 1999) e Emmons (1990) realizaram compilações ilustradas atualizando a visão de conjunto das espécies. Anderson (1982 e 1997), Lemos, Weksler & Bonvicino (2000), Voss, Lunde & Simmons (2001) e Vargas, Tarifa & Cortez (2002) também elucidaram pontos específicos sobre a sistemática do gênero, e mais recentemente, Solari (2003) descreveu M. ronaldi do leste do Peru, e em 2007 M. handleyi.
Em 1991 realizei como dissertação de mestrado uma revisão do gênero Monodelphis, que não foi publicada, mas acabou tornando-se conhecida por ser a única referência atualizada naquela época e pela divulgação que realizei posteriormente em CD-rom enviando para museus europeus e norte americanos, o que certamente motivou discussões sobre o assunto a partir de então.
Este trabalho é uma continuidade dessa revisão, que agora é apresentada de forma mais ampla, tendo como objetivo, fornecer uma classificação atualizada das espécies do gênero Monodelphis, além de situá-lo no contexto dos Marsupiais como um todo.
As 22 espécies reconhecidas aqui estão reunidas em seis grupos de espécies, construídos com base em uma análise filogenética amadurecida por décadas de pesquisa incessante. Para isso utilizei o material depositado em 3 museus brasileiros, Museu de Zoologia da USP (MZUSP), Museu Nacional do Rio de Janeiro (MNRJ) e Museu Paraense Emílio Goeldi (MPEG). As espécies M. osgoodi e M. reigi, que não estão presentes em nossas coleções, são comentadas e comparadas com base na literatura.
Realizei também uma análise estatística aproveitando que a espécie M. domestica está representada por uma grande quantidade de espécimes (perto de mil exemplares) nas coleções do MNRJ (todos coletados no nordeste brasileiro pelo Serviço da Peste). Essa análise forneceu dados sobre a variação geográfica, etária e sexual de M. domestica, que foram utilizados na comparação e compreensão das outras espécies do gênero por extrapolação.
Nesta revisão apresento descrições acuradas de cada uma das espécie, chaves de identificação das mesmas, mapas de distribuição geográfica, proponho uma filogenia para os principais grupos de marsupiais permitindo uma visão relativa da posição do gênero Monodelphis, num contexto mais amplo e para complementar o trabalho realizo também, um estudo da biogeografia do gênero.
MATERIAL E MÉTODOS
Coleções de Museus - Neste estudo foram utilizados exemplares depositados nos três principais museus brasileiros: Museu Nacional do Rio de Janeiro (MNRJ), (cerca de mil e duzentos exemplares), Museu Paraense Emilio Goeldi (MPEG), (97 exemplares) e Museu de Zoologia da USP (MZUSP), (242 exemplares).
A maior parte do material examinado consistiu de peles cheias acompanhadas de seus respectivos crânios. Uma pequena parte desse material estava em meio líqüido (conservação total de exemplares em álcool ou formol).
Estudo do material conservado em meio líquido - Esse material permitiu um estudo de caracteres, que de um modo geral, não foram levados em conta pelos sistematas que trabalharam com os didelfídeos até então. Tais caracteres são: rinário, orelha, patas e cauda. Cada um deles foi descrito de forma comparativa. Algumas peles secas foram colocadas em câmara úmida para permitir a análise desses caracteres.
Estudos da pelagem - A pelagem é descrita com base nas peles secas. Este caráter é descrito para todo o gênero com uma caracterização do tamanho dos pêlos nas diferentes partes do corpo e dentro de cada grupo de espécies são fornecidos os padrões gerais de coloração. Nas espécies a pelagem é descrita primeiro na parte dorsal, seguida das laterais, parte ventral, patas e cauda. Quando necessário o padrão de coloração das bandas dos pêlos é fornecido.
Morfologia craniana - O crânio é descrito apenas na introdução de cada grupo de espécies com comparações entre espécies do mesmo grupo ou de grupos morfologicamente próximos. A monenclatura foi baseada em Flower, (1966).
Morfologia dentária - Os dentes são comparados inicialmente como um todo dentro do gênero e depois tratados mais detalhadamente nas introduções dos grupos de espécies. Foram feitas comparações de tamanho e proporcões entre as espécies e características principais tais como proporções das cristas, estilos e estílidos. As ilustrações dos crânios e dentes permitem um acompanhamento visual. A monenclatura foi baseada em Hershkovitz, (1959).
Medidas corporais - Fornecidas nas etiquetas das peles foram agrupadas e comparadas, tanto em valor absoluto quanto com relação a suas proporções.
Medidas cranianas - Foram realizadas vinte e seis medidas cranianas por exemplar:
1. Comprimento do crânio (côndilo-basal).
2. Comprimento palatal (distância entre a parte anterior da pré-maxila e a parte posterior do palatino).
3. Comprimento da fossa órbito-temporal (medido inferiormente, da parte posterior do maxilar até a borda do alisfenóide).
4. Comprimento I1-M4 (distância entre a borda bucal do incisivo 1 até a borda distal do molar 4 superior).
5. Comprimento dos nasais (maior comprimento dos nasais).
6. Comprimento da mandíbula (distância entre a borda bucal do incisivo 1 até a borda posterior do côndilo mandibular).
7. Comprimento i1-m4 (distância entre a borda bucal do incisivo 1 até a borda distal do molar 4 inferiores).
8. Largura C-C externa (distância entre as faces bucais dos caninos superiores, tomada próxima às raízes).
9. Largura P2-P2 externa (maior distância entre as faces bucais dos pré-molares superiores).
10. Largura posterior do focinho (distância entre as bordas dos forâmens infra-orbitais).
11. Largura M3-M3 externa (maior distância entre as bordas bucais dos molares superiores).
12. Largura dos nasais (maior largura dos nasais conjuntamente).
13. Largura zigomática (maior largura externa entre os arcos zigomáticos).
14. Largura da caixa craniana (distância entre partes mais posteriores das fossas glenóides).
15. Largura entre os processos timpânicos (maior distância entre os processos timpânicos dos perióticos).
16. Largura entre os processos parocipitais (maior distância externa).
17. Altura da caixa craniana (medida da parte superior e mais posterior da crista sagital até o centro do basocipital posterior).
18. Altura do processo coronóide (distância entre a extremidade dorsal do processo coronóide até a borda ventral da fossa do masseter).
19. Constrição pós-orbital (distância entre as bordas laterais internas da constrição).
20. Comprimento M1-M3 (distância entre a borda mesial do cíngulo bucal do molar superior 1 até a borda distal do cíngulo bucal do molar superior 3).
21. Comprimento M1-M4 (distância entre a borda mesial do cíngulo bucal do molar superior até a borda distal do cíngulo bucal do molar superior 4).
22. Comprimento m1-m4 (distância entre a borda mesial do cíngulo lingual do molar inferior 1 até a borda distal do cíngulo lingual do molar inferior 4).
23. Largura do M4 esquerdo (maior distância entre as bordas lingual e bucal do molar superior esquerdo).
24. Largura do M4 direito (maior distância entre as bordas lingual e bucal do molar superior direito).
25. Largura do P2 esquerdo (maior distância entre as bordas do pré-molar superior esquerdo).
26. Largura do P2 direito (maior distância entre as bordas do pré-molar superior direito).
Essas medidas foram realizadas sob lupa estereoscópica, com o auxilio de um paquímetro de precisão igual a 0,01 mm. A figura 1 mostra os pontos de referência utilizados nas mensurações.

Foram calculadas médias, desvios padrões, variâncias e coeficientes de variação das medidas realizadas nos exemplares de cada uma das espécies. Devido ao grande número de exemplares disponíveis, em M. domestica foi feito um estudo de variação geográfica. As localidades amostradas são fornecidas na análise.
As medidas foram divididas em dois grupos: medidas variáveis com o crescimento do animal e medidas que não apresentam variação ao longo da vida.
Foi realizado o teste de análise de variância (ANOVA) modelo II e também análises de comparações múltiplas, testes Student-Newman-Keuls(SNK) e Turkey, nas medidas do segundo grupo.
As medidas foram divididas em dois grupos: medidas variáveis com o crescimento do animal e medidas que não apresentam variação ao longo da vida.
Foi realizado o teste de análise de variância (ANOVA) modelo II e também análises de comparações múltiplas, testes Student-Newman-Keuls(SNK) e Turkey, nas medidas do segundo grupo.
As medidas restantes foram analisadas da seguinte forma: Uma vez selecionados todos os indivíduos adultos (que são aqueles que apresentaram exclusivamente dentição permanente), foram separadas dentro desta amostra as medidas extremas. Por exemplo: o maior e o menor dos comprimentos dos crânios. Com base nestas duas medidas extremas foi calculada a porcentagem de crescimento durante a vida adulta. Todas as medidas variáveis com o crescimento foram assim tratadas (tabelas 1-10). Usei tabém o programa Excel para construir tabelas, e posteriormente produzir gráficos de crescimento, através de curvas logarítmicas, usando os coeficientes angulares na comparação das curvas.
Sistemática
Esta revisão sistemática obedeceu as seguintes etapas:
1. Separação das formas existentes no material examinado, sem se preocupar com nomes ou qualquer identificação prévia.
2. Observação da consistência dos grupos separados: possíveis variações ou continuidades de caracteres entre grupos.
3. Observação de variações etária e sexual.
4. Observação de possíveis variações geográficas nas continuidades de caracteres existentes.
5. Relação entre as forma e os nomes existentes (identificação), nesse caso é imprescindível o uso das descrições originais das espécies para que erros de classificação não se perpetuem.
6. Organização da sinonímia das espécies para determinação dos sinônimos "junior" e sênior de acordo com o Código Internacional de Nomenclatura Zoológica.
7. Descrição de espécies novas.
8. Ordenação das espécies segundo critérios filogenéticos (grupos monofiléticos).
O reconhecimento de espécies a partir de exemplares carrega consigo algumas limitações, o tamanho e a representatividade (geográfica, etária ou sexual) da amostra. Por isso, as conclusões obtidas em qualquer trabalho desse tipo sempre estarão condicionadas a esses fatores.
Estabelecer limites entre espécies nem sempre é uma tarefa simples, às vezes dados geográficos são levados em conta ou mesmo extrapolações a partir de certos padrões já observados em outras espécies.
7. Descrição de espécies novas.
8. Ordenação das espécies segundo critérios filogenéticos (grupos monofiléticos).
O reconhecimento de espécies a partir de exemplares carrega consigo algumas limitações, o tamanho e a representatividade (geográfica, etária ou sexual) da amostra. Por isso, as conclusões obtidas em qualquer trabalho desse tipo sempre estarão condicionadas a esses fatores.
Estabelecer limites entre espécies nem sempre é uma tarefa simples, às vezes dados geográficos são levados em conta ou mesmo extrapolações a partir de certos padrões já observados em outras espécies.
Filogenia
Os pontos centrais dessa análise são:
1. Levantamento de sinapomorfias – Através da comparação com grupos externos (no caso os outros marsupiais e os Insectivora) decidiu-se as condições dos caracteres (sinapomorfias e simplesiomorfias). As sinapomorfias são utilizadas para reunir as espécies em grupos monofiléticos. Na análise das inter-relações no nível de espécie o estabelecimento de séries de transformações foram de grande importância devido a sutileza das diferenças observadas.
2. A regra da parcinômia foi aplicada a seguir para resolver os problemas de hipóteses conflitantes.
Problemas na análise das interrelações de espécies muito próximas:
a) Heterobatmia entre espécies irmãs nem sempre é possível, pois devemos considerar a possibilidade da espécie ancestral continuar viva, ao lado de sua irmã mais derivada (com suas autapomorfias). Análises entre espécies muito próximas freqüentemente se deparam com tal situação.
b) É perfeitamente possível realizarmos uma análise cladística entre espécies irmãs (duas espécies originadas de um ancestral exclusivo), subespécies, raças ou formas geográficas. Para isso devemos trata-las a priori como se fossem espécies (como módulos isolados), pois uma das espécies pode ser mais intimamente relacionada a uma determinada raça de uma espécie irmã e não a espécie como um todo. A compreensão dessas íntimas relações é de grande importância em uma análise biogeográfica baseada em um cladograma. A representação de um cladograma resultante de uma análise desse tipo deverá ser discutida mais adiante, por apresentar uma visão aparentemente conflitante com a lógica estabelecida por Hennig (1950).
Biogeografia
A distribuição das espécies, os tipos de ambientes ocupados pelas espécies do gênero, os casos de simpatria, parapatria e os processos de especiação e evolução relacionados a biogeografia são tratados em conjunto na biogeografia.
Partindo da construção de mapas de distribuição das espécies, tentei relacionar essas distribuições com as principais formações vegetais associadas.
A análise biogeográfica tem por base os inúmeros trabalhos já realizados na área de biogeografia. Entre eles chamo a atenção para Croizat (1974), Raven & Axelrod (1974) e Rizzini (1979).
Calcada principalmente nos mapas de distribuição geográfica das espécies, esta análise leva em conta igualmente o possível ambiente ocupado pelas espécies nessas áreas de distribuição. Procura também determinar na medida do possível os eventos responsáveis pela irradiação do grupo .
Método de análise biogeográfica
A análise obedece a dois postulados principais:
1. A base do primeiro postulado: As modificações genéticas ocorrem como respostas às modificações ambientais (Stebbin, apud Rizzini, 1979: 339), ou seja, uma espécie muda quando o ambiente ao seu redor também está mudando. Se tivermos dois ambientes diferentes, ocupados por duas espécies irmãs, a mais plesiomórfica deve estar no ambiente menos modificado.
2. A base do segundo postulado: A especiação geralmente acontece por isolamento espacial de populações de uma mesma espécie, e sendo assim "se um modo de especiação alopátrida é postulado, então a simpatria dentro de um grupo monofilético é evidência de dispersão" (Croizat, 1974, apud Wiley, 1981: 303).
Este modelo serve para explicar especiação e dispersão de pequenos mamíferos terrestres e não pretende extrapolar esse âmbito.
As localidades dos exemplares examinados foram plotadas em mapas e o padrão encontrado foi relacionado à distribuição das grandes formações vegetais da América do Sul.
1. A base do primeiro postulado: As modificações genéticas ocorrem como respostas às modificações ambientais (Stebbin, apud Rizzini, 1979: 339), ou seja, uma espécie muda quando o ambiente ao seu redor também está mudando. Se tivermos dois ambientes diferentes, ocupados por duas espécies irmãs, a mais plesiomórfica deve estar no ambiente menos modificado.
2. A base do segundo postulado: A especiação geralmente acontece por isolamento espacial de populações de uma mesma espécie, e sendo assim "se um modo de especiação alopátrida é postulado, então a simpatria dentro de um grupo monofilético é evidência de dispersão" (Croizat, 1974, apud Wiley, 1981: 303).
Este modelo serve para explicar especiação e dispersão de pequenos mamíferos terrestres e não pretende extrapolar esse âmbito.
As localidades dos exemplares examinados foram plotadas em mapas e o padrão encontrado foi relacionado à distribuição das grandes formações vegetais da América do Sul.
Elaboração dos mapas de distribuição geográfica.
Os mapas de distribuição geográfica foram modificados a partir de Hueck, K., & P. Seibert. 1972. Foi dada uma evidência entre as regiões florestadas (representadas em verde) e as vegetações abertas (em amarelo claro) que incluem cerrado e caatinga. O mapa foi todo clareado para evidenciar os pontos plotados.
O mapa das localidades amostradas de Monodelphis domestica do nordeste brasileiro foi modificado a partir de um mapa retirado do Atlas Encarta Microsoft.
Os mapas da América do Sul usados no estudo da evolução do gênero, parte de biogeografia, foram construidos seguindo três orientações:
1 -
2 -
3 - Divisores de águas ou limites entre as bacias fluviais. Se os rios forem encarados como barreiras para a dispersão da fauna terrestre não-natante e não-volante. Então, os corredores preferenciais para dispersão da fauna terrestre seriam os divisores de águas situados entre as bacias fluviais. As áreas do Brasil central, onde a pluviosidade é mais intensa, onde existem [ou existiram] inúmeras manchas de matas e também onde se localizam os divisores de águas, foram adotadas como o possível corredor de ligação entre a Amazônia e a Mata Atlântica.
Gênero Monodelphis Burnett, 1830
Sorex Muller (1776: 36), parte.
Didelphis Erxleben (1777: 80), parte.
Didelphys Schreber (1778: 548) parte.
Viverra Shaw (1800: 432), parte.
Mustela Bechstein (1800: 354), parte.
Philander Tiedemann (1808: 427), parte.
Sarigua Muirhead (1819: 429), parte.
Monodelphis Burnett (1830: 351).
Peramys Lesson (1842: 187).
Micoureus Lesson (1842: 187).
Grymaeomys Burmeister, 1854.
Grymaeomys Burmeister, 1854.
Hemiurus Gervais (1855-1856: 101)
Microdelphys Burmeister (1856: 83), parte.
Monodelphiops Matschie (1916: 260).
Minuania Cabrera (1919: 43).
A lista acima foi modificada a partir de Cabrera, 1958.
Espécie-tipo: M. brachyura Burnett = Didelphis brevicaudata Erxleben, 1777.
A primeira espécie conhecida deste gênero foi descrita como Sorex americanus por Muller (1776: 36), baseada em "La Musaraigne du Brèzil" de Buffon, 1767:160 (que por sua vez foi totalmente baseada no Mus araneus de Marcgrave, 1648: 229).
A segunda espécie do gênero foi descrita por Erxleben (1777: 80). Ele a descreveu como Didelphis brevicaudata, baseado no Muris sylvestris americani foemina de Seba, numa época na qual todos os marsupiais conhecidos eram colocados no gênero Didelphis.
Shaw (1800: 432) baseado em Le Touan de Buffon, considerada por este como um furão, descreveu Viverra touan. Bechstein (1800: 354) com base na mesma referência descreveu Mustela touan.
Tiedemann (1808: 427) propõe Philander brachyurus para Didelphys brachyuros Schreber. Muirhead (1830: 429) usa Sarigua para D. brachyuros Schreber na "Brewster’s Edinburgh Encyclopaedia". Burnett (1830: 351) propõe o gênero Monodelphis para "D. dorsigerens" e D. brachyura. "D. dorsigerens" supostamente representa uma corrupção de D. dorsigera L. considerada pelo menos em parte, como um sinônimo junior de Marmosa murina L., por isso Monodelphis fica associado à Didelphys brachyuros Schreber (= M. brevicaudata Erxleben). Lesson (1842: 187) propõe Peramys com P. brachyura e P. tristriata e Micoureus com M. tricolor, para as espécies de Monodelphis até então conhecidas.
Gervais (1855: 101) propõe Hemiurus para Didelphys hunteri (Waterhouse), incluindo (Didelphys tristiata [sic], tricolor et brachyura des auteurs)". Burmeister (1856: 83) coloca as espécies D. tristriata, D. tricolor, D. brachyura, D. domestica e D. unistriata no sub-gênero Microdelphys, que aparece também como simples binômio. Matschie (1916: 260) propõe Monodelphiops para Microdelphys sorex (Hensel). Cabrera (1919: 43) cria o gênero Minuania para Didelphys dimidiata (Wagner)
Alguns nomes foram usados como subgênero:
Peramys foi subgênero de Didelphis em: Didelphys (Peramys) henseli: Thomas (1888a: 159). Thomas (1888b 360).
Minuania em Monodelphis (Minuania) dimidiata: Cabrera (1958: 10) e Monodelphis em: Monodelphis (Monodelphis) brevicaudata brevicaudata: Cabrera (1958: 7).
De acordo com meus resultados filogenéticos esses subgêneros não são satisfatórios. Minuania concorda em parte com o grupo monofilético formado por M. dimidiata e M. brevicaudis, porém em contraposição a [Monodelphis] como subgênero não satisfaz a condição de monofilia, pois o subgênero Monodelphis ficaria parafilético. Separei o gênero em 6 grupos de espécies, pois não vejo necessidade de se criar subgêneros, só iria complicar as citações desnecessáriamente.
Tamanho pequeno, com um comprimento máximo em torno de 28 cm. Cauda curta, pouco mais comprida que a metade do comprimento do corpo, com escamas, pouco pilosa e muito pouco preênsil. Orelhas nuas, curtas e arredondadas, pelo curto, sem marsúpio. Crânio geralmente largo, oval e achatado na parte superior. Palato sem o segundo par de forâmens. Os termos : pequeno, curto, largo e pouco, são termos relativos e se referem, por comparação, aos demais marsupais sul-americanos.
O gênero Monodelphis ocupa, na sua área de distribuição, o mesmo nicho ocupado pelos insetívoros terrestres de outras regiões do planeta.
Suas características, como cauda curta, orelhas pequenas, aliadas ao reduzido diâmetro dos olhos e hálux pouco oponível em relação aos outros artelhos, mostram uma adaptação semi-fossorial.
Distribuição geográfica - Ocorre no Panamá, Colômbia, Venezuela, Guianas, Equador, Peru, Bolívia, Brasil, Paraguai e na Argentina até Mar del Plata.
Habitats
Vivem em florestas úmidas, no Cerrado, campos e em regiões semi-áridas como a Caatinga e o Chaco.
Vivem em florestas úmidas, no Cerrado, campos e em regiões semi-áridas como a Caatinga e o Chaco.
Organização da Pelagem
A pelagem das espécies do gênero Monodelphis não difere muito da pelagem das espécies menores da família Didelphidae, de modo geral os pelos são curtos. Os pelos variam de tamanho e forma nas várias regiões do corpo. A cauda, os dedos e os artelhos apresentam um revestimento de escamas e a cada escama estão geralmente associados três pelos (fig. 16). Esse grupo de pelos recebe o nome de tríade e geralmente o pêlo central da tríade é mais longo e grosso que os laterais.

Os pelos que se encontram associados às escamas geralmente são os mais curtos de toda pelagem. Existem pelos curtos como o das tríades na ponta do focinho e nas orelhas. No resto da cabeça, os pelos mais curtos se encontram nas partes laterais do focinho, ao redor dos olhos e da boca. Da ponta do focinho até o meio do corpo, os pelos são gradativamente mais longos. Na cabeça, os pelos são especialmente longos na frente das orelhas; no pescoço, são mais longos lateralmente. Na parte inferior do pescoço, junto do peito, existe uma área estreita central de pelos muito curtos, aos quais, segundo Beddard (1888: 353), está associada uma glândula (fig. 2). Nas regiões dorsal e lateral do corpo os pelos são mais longos na parte central, daí até a base da cauda são gradativamente mais curtos; ventralmente são mais longos na região inguinal. As patas são revestidas dorsalmente por pelos curtos como os da cabeça e ventralmente não possuem pelos. Nas partes internas dos braços e das pernas os pelos são curtos e escassos; externamente são mais curtos próximo às patas e gradativamente mais longos em direção ao corpo (fig. 2).
Os pelos curtos possuem uma base fina e curta, seguida de um engrossamento, estreitando-se novamente até terminar em ponta bem fina.
Os pelos mais longos, que revestem o pescoço, o corpo, os braços e as pernas, são de três tipos diferentes: aristiformes, setiformes e lanugem. Os aristiformes e setiformes são finos na base continuando com a mesma espessura até aproximadamente a metade do comprimento, então se engrossando e novamente afinando até a ponta; a lanugem tem espessura uniforme só afinando na ponta.
Os aristiformes são mais longos e grossos que os demais e se sobressaem na pelagem, os setiformes são de grossura e comprimento médios e a lanugem por sua vez é mais fina e curta.
Além desses pêlos, muito numerosos em algumas regiões do corpo, existem pelos que podem ser bem mais longos e grossos que os aristiformes e que desempenham papel sensitivo, esses pelos recebem o nome de vibrissas.
As vibrissas podem ser classificadas em oito tipos: (1) Mistaciais, agrupadas em duas áreas separadas, uma em cada lado do focinho; em cada uma dessas áreas essas vibrissas se organizam em cinco fileiras longitudinais principais, que por sua vez se dispõem no sentido dorso-ventral. Podem chegar a trinta e dois pares no total. (2) Supraorbitais, localizadas logo acima de cada olho, geralmente dois pares. (3) Suborbitais, localizadas nas bochechas e dispostas no sentido dorsoventral, geralmente oito pares. (4) Mentonianas, localizadas nas bordas anteriores dos lábios inferiores, geralmente três pares. (5) Inter-ramais, localizadas em uma área única central na garganta, geralmente cinco unidades. (6) Ulnar-carpais, localizadas na parte externa dos pulsos, geralmente três pares. (7) Antebraquiais, localizadas posteriormente às ulnar-carpais no antebraço, geralmente um par. (8) Anconeais, localizadas na porção média do antebraço externamente, posteriormente as ulnar-carpais, geralmente um par (fig. 3).

Nas regiões dorsais, laterais e ventrais os pelos podem ser de uma única cor ou apresentar bandas com diferentes cores. As bandas variam de duas a quatro.
Geralmente os pelos possuem uma banda basal cinzenta, que nos aristiformes e setiformes corresponde à porção compreendida entre a raiz do pelo e a região de engrossamento.
Os aristiformes dorsais e laterais podem apresentar de duas a quatro bandas, geralmente duas, uma banda basal cinza e uma terminal larga, preta ou marrom. Os aristiformes ventrais podem ser de uma única cor ou apresentar duas ou três bandas; nesse caso, sempre a banda basal é cinza.
Os setiformes dorsais e laterais podem apresentar de duas a quatro bandas. Geralmente são quatro: uma banda basal cinza, uma mediana preta ou marrom, uma subterminal que pode variar entre branca amarelada, amarela e laranja, e uma banda terminal pequena, preta ou marrom. Os setiformes ventrais podem ser de uma única cor ou apresentar duas ou três bandas, nesse caso com a banda basal sempre cinza.
A lanugem de modo geral é uniformemente cinza ou marrom, de tonalidade clara ou escura.
As vibrissas normalmente possuem cor uniforme, podendo ser pretas ou brancas, dependendo de sua localização. Geralmente as dorsais são escuras e as ventrais claras.

Caracteres cranianos dos jovens e classificação etária dos crânios

Dentição





Anatomia externa







Pelagem: padrões de cor.
Os padrões de cor encontrados nos exemplares examinados podem ser colocados em quatro grupos:
Os padrões de cor encontrados nos exemplares examinados podem ser colocados em quatro grupos:
1-Coloração dorsal cinza-grisalho e partes laterais variando de cinza-amarelado até marrom bem avermelhado.
2-Coloração dorsal cinza-grisalho com a cabeça marrom alaranjada e traseiro marrom avermelhado.
3-Coloração dorsal marrom amarelada com traseiro marrom avermelhado e três faixas escuras longitudinais.
4-Coloração dorso-lateral uniforme marrom.
Caracteres cranianos adultos do gênero Monodelphis
O maior comprimento do crânio nos adultos varia entre 28 mm em M. iheringi a 47 mm em M. domestica; forma do crânio oval, achatada dorsalmente (figs. 20A-20C); cristas temporais levemente marcadas ou unidas numa crista sagital; perfil nasal de reto a convexo; nasais alongados, com um alargamento nos dois quintos proximais, geralmente terminado em ponta; pré-maxilar com processo ascendente longo ou médio, paralelo ao nasal, forâmem infraorbital situado acima do P3 ou M1; órbita de um quinto a um sétimo do comprimento do crânio; jugal dobrado na região orbital, com ângulo interno da borda lateral variando entre 160o e 178o, lacrimal com porção anterior côncava ou convexa; esfenóide alargado, interpondo-se amplamente entre o frontal e o forâmen orbitosfenóide ou estreito e afilado na porção superior, interpondo-se muito pouco; processos pós-orbitais entre pouco pronunciados ou ausentes, próximos da constrição intertemporal, ou levemente afastados; comprimento da caixa craniana entre um terço a quase metade do comprimento do crânio; supraocipital com cristas bem desenvolvidas ou rudimentares na porção mediana; palato pouco mais comprido que a metade do comprimento do crânio, arcada em forma de "V", com séries C-P3 paralelas ou não, as séries M1-M4 de comprimento igual ou menor que as séries C-P3; forâmens palatais posteriores longos, de P3 a M3 ou curtos, de M1 a M3; alisfenóides com crista transversal presente ou ausente; foramen rotundum tão próximo do canal transverso quanto o forâmen da carótida ou até três vezes mais afastados que este; forâmens das carótidas tão próximos entre si quanto cada um deles dos canais transversos adjacentes, ou mais afastados entre si que destes; bula auditiva com processo alisfenóide desenvolvido e avançado em direção ao periótico ventralmente ou mais curto, deixando um espaço meso-ventral maior entre este e o periótico; processo timpânico do periótico em ponta, ou arredondado, variando entre alongado e aproximadamente quadrado; foramen magnum entre arredondado e achatado no sentido dorso-ventral; perfil da borda inferior do ramo horizontal da mandíbula levemente curvo ou aproximadamente reto; ramo ascendente inclinado na direção posterior, com ângulo interno entre 100o e 200o, processo coronóide estreito, mais ou menos triangular, ou alargado; distância entre a projeção externa da borda inferior da fossa massetérica e o processo condilar variável; sínfise mandibular com comprimento entre um quarto a quase um terço do comprimento da mandíbula; inclinação interna do processo angular entre 90o e 130o (Figs. 4, 20A-20D).
O maior comprimento do crânio nos adultos varia entre 28 mm em M. iheringi a 47 mm em M. domestica; forma do crânio oval, achatada dorsalmente (figs. 20A-20C); cristas temporais levemente marcadas ou unidas numa crista sagital; perfil nasal de reto a convexo; nasais alongados, com um alargamento nos dois quintos proximais, geralmente terminado em ponta; pré-maxilar com processo ascendente longo ou médio, paralelo ao nasal, forâmem infraorbital situado acima do P3 ou M1; órbita de um quinto a um sétimo do comprimento do crânio; jugal dobrado na região orbital, com ângulo interno da borda lateral variando entre 160o e 178o, lacrimal com porção anterior côncava ou convexa; esfenóide alargado, interpondo-se amplamente entre o frontal e o forâmen orbitosfenóide ou estreito e afilado na porção superior, interpondo-se muito pouco; processos pós-orbitais entre pouco pronunciados ou ausentes, próximos da constrição intertemporal, ou levemente afastados; comprimento da caixa craniana entre um terço a quase metade do comprimento do crânio; supraocipital com cristas bem desenvolvidas ou rudimentares na porção mediana; palato pouco mais comprido que a metade do comprimento do crânio, arcada em forma de "V", com séries C-P3 paralelas ou não, as séries M1-M4 de comprimento igual ou menor que as séries C-P3; forâmens palatais posteriores longos, de P3 a M3 ou curtos, de M1 a M3; alisfenóides com crista transversal presente ou ausente; foramen rotundum tão próximo do canal transverso quanto o forâmen da carótida ou até três vezes mais afastados que este; forâmens das carótidas tão próximos entre si quanto cada um deles dos canais transversos adjacentes, ou mais afastados entre si que destes; bula auditiva com processo alisfenóide desenvolvido e avançado em direção ao periótico ventralmente ou mais curto, deixando um espaço meso-ventral maior entre este e o periótico; processo timpânico do periótico em ponta, ou arredondado, variando entre alongado e aproximadamente quadrado; foramen magnum entre arredondado e achatado no sentido dorso-ventral; perfil da borda inferior do ramo horizontal da mandíbula levemente curvo ou aproximadamente reto; ramo ascendente inclinado na direção posterior, com ângulo interno entre 100o e 200o, processo coronóide estreito, mais ou menos triangular, ou alargado; distância entre a projeção externa da borda inferior da fossa massetérica e o processo condilar variável; sínfise mandibular com comprimento entre um quarto a quase um terço do comprimento da mandíbula; inclinação interna do processo angular entre 90o e 130o (Figs. 4, 20A-20D).
De modo geral os jovens apresentam o focinho mais curto que os adultos, mas é a dentição a característica mais importante na diagnose dos jovens.
Nos marsupiais o terceiro pré-molar sofre uma substituição. Os dentes decíduos molariformes precedem tanto os pré-molares superiores quanto os inferiores.
A classificação etária usada nas Tabelas apresentadas no final do trabalho, foi a seguinte:
1. Jovem: quando o exemplar apresentar pelo menos um pré-molar decíduo (fig. 5).

2. Subadulto (adulto jovem): quando o exemplar apresentar pelo menos um dos terceiros pré-molares em erupção, com mesoestilo menos projetado que o dististilo do segundo pré-molar adjacente (fig. 6).

2. Subadulto (adulto jovem): quando o exemplar apresentar pelo menos um dos terceiros pré-molares em erupção, com mesoestilo menos projetado que o dististilo do segundo pré-molar adjacente (fig. 6).

3. Adulto - quando o exemplar apresentar o mesoestilo do terceiro pré-molar mais projetado que o dististilo do segundo pré-molar adjacente (fig. 6).
4. Adulto velho - quando o exemplar apresentar os molares desgastados, os caninos bem projetados e o palato posterior bem avançado posteriormente.
Fórmula dentária : i 5/4 c 1/1 p 3/3 m 4/4

Incisivos - os menores dentes do sistema dentário.
superiores
I1 - cônico, com curvatura no sentido mesial, levemente proodonte ou opistodonte, separado dos demais incisivos por um pequeno diástema.
I2-I5 - tridentados; eocone, mesoestilo e dististilo presentes; opistodontes; paracrista aproximadamente igual a pós-metacrista; cíngulo bucal pouco maior que o cíngulo lingual.
I2- menor que os demais.
I3 e I4- quase iguais entre si.
I5- maior ou igual a I3 e I4. (fig. 7)
inferiores - maiores que os superiores, alongados e espatulados; semelhantes entre si, com pequena redução de tamanho no sentido distal; cíngulo lingual bem desenvolvido, mais ou menos triangular em vista oclusal; i2 deslocado, com implantação mais interna, como na maioria dos marsupiais com exceção de Dromiciops; marcadamente proodontes (fig. 7)
Caninos - os maiores dentes do sistema dentário, de crescimento contínuo, mais ou menos cônicos, com um achatamento buco-lingual às vezes bem pronunciado; algumas vezes mais desenvolvidos nos machos; os superiores proodontes ou ortodontes; os inferiores proodontes; nos jovens a ponta ainda possui uma semelhança com os pré-molares, com mesoestilos e dististilos presentes; superiores mais robustos que os inferiores nos adultos.
Pré-molares - sagitados, tridentados (eocone desenvolvido, mesoestilo e dististilo presentes), achatados no sentido buco-lingual; pós-metacristas mais aguçadas, côncavas; paracristas arredondadas, convexas.
Superiores
P1 - menor que os demais pré-molares; comprimento meso-distal dois terços de P3, altura da coroa metade de P3; secção transversal triangular; comprimento meso-distal quase o dobro da altura da coroa; cíngulos bucal anterior e posterior quase em linha reta; cíngulos lingual anterior e posterior em ângulo obtuso; cíngulo lingual posterior bem desenvolvido; raízes expostas nos adultos, a anterior as vezes mais exposta e mais fina que a posterior; pós-metacrista duas vezes mais longa que a paracrista; dististilo geralmente mais desenvolvido que o mesoestilo; eixo da coroa com inclinação anterior.
P2 - altura da coroa pouco menor que em P3; comprimento meso-distal aproximadamente igual a P3; secção transversal em losango; altura da coroa pouco menor que o comprimento meso-distal; cíngulo bucal anterior e posterior em ângulo obtuso; cíngulo lingual anterior e posterior igualmente; cíngulos bucal e lingual posteriores desenvolvidos, principalmente o bucal; raízes como em P1; pós-metacrista igual ou um pouco mais longa que a paracrista; dististilo e mesoestilo desenvolvidos; eixo do dente com inclinação posterior.
P3 - altura da coroa aproximadamente igual ao comprimento meso-distal; secção transversal oval; cíngulos lingual e bucal aproximadamente curvos; cíngulos bucal e lingual posteriores menos desenvolvidos que em P2; raízes bem expostas nos adultos, principalmente a anterior; pós-metacrista igual ou um pouco mais curta que a paracrista; mesoestilo e dististilo menos desenvolvidos que em P2; eixo dos dentes com inclinação posterior (fig. 8).

Inferiores
p1 - menor que os demais pré-molares; comprimento meso-distal igual ou dois terços de p3; secção transversal aproximadamente em losango; comprimento meso-distal entre o dobro e o triplo da altura da coroa; cíngulos bucal anterior e posterior igualmente; cíngulo lingual posterior bem desenvolvido, geralmente duas vezes mais longo que o anterior; raízes expostas nos adultos, a anterior mais exposta que a posterior; pós-metacrista duas vezes mais longa que a parametacrista; dististilo mais desenvolvido que o mesoestilo; eixo da coroa com inclinação anterior.
p2 - altura da coroa pouco menor que em p3; comprimento meso-distal maior que em p3; secção transversal entre oval e triangular; altura da coroa entre metade ou três quartos do comprimento meso-distal; cíngulos bucal anterior e posterior, cíngulos lingual anterior e posterior em ângulo obtuso; cíngulos bucal e lingual posteriores desenvolvidos; raízes como em p1; paracrista entre dois terços e metade do comprimento da pós-metacrista; dististilo desenvolvido; mesoestilo reduzido; eixo do dente com inclinação anterior ou perpendicular ao ramo mandibular.
p3 - o maior pré-molar; altura da coroa igual ou menor que o comprimento meso-distal; secção transversal oval; cíngulos bucal e lingual em curva; cíngulos bucal e lingual posteriores desenvolvidos, principalmente o bucal; raízes bem expostas nos adultos mais que nos outros pré-molares, com altura da área exposta, as vezes maior que a altura da coroa; paracrista variando entre pouco menor e aproximadamente metade da pós-metacrista; dististilo desenvolvido; mesoestilo reduzido; eixo do dente com inclinação entre perpendicular e posterior ao ramo da mandíbula (fig. 8),
Molares
Molares

Molares superiores M1, M2 e M3 semelhantes, dilambdomórficos, metacone sempre presente, freqüentemente maior que o eocone; eocrista em forma de "W"; cíngulo bucal sempre expandido em uma prateleira larga, subdividida por uma dobra distinta em área paraestilar e metaestilar, cada uma com três ou quatro ectaestilos; cíngulo lingual sobre uma base triangular onde se localiza o protocone; triangulares em secção transversal. M4 comprimido no sentido antero-posterior do palato (fig. 9 e 10).

M1 - menor que M3 e M4, comprimento meso-distal da coroa aproximadamente como em P3 ou pouco maior; comprimento meso-dististilo pouco maior que a largura mesoestilo-protocone; bacia do cíngulo paraestilar igual ou pouco menor em área que a bacia do trígono; bacia do cíngulo metaestilar duas ou três vezes maior em área que a bacia do cíngulo paraestilar; paracrista entre metade e um terço do comprimento da pós-metacrista; pós-paracrista pouco mais longa que a metade da pré-metacrista; protocrista aproximada-mente igual ou pouco maior que a plagiocrista em comprimento; crista do paraestilo pouco mais longa que a metade da crista metaestilar; cíngulo bucal bem mais desenvolvido que o cíngulo lingual; mesoestilo e dististilo muito reduzidos, ectoestilo l bem desenvolvido, maior ou igual ao eocone; ectoestilo j menor que l, porém mais desenvolvido que os demais estilos; ectoestilo m e k quando presentes muito reduzidos.
M2 - menor que M3; comprimento meso-distal da coroa aproximadamente igual a M1; comprimento meso-dististilo igual ou pouco menor que a largura mesoestilo-protocone; bacia do cíngulo paraestilar aproximadamente igual à bacia do trígono, maior que M1 em área; bacia do cíngulo metaestilar aproximada-mente o dobro da bacia do cíngulo metaestilar; paracrista aproximadamente dois terços da pós-metacrista; pós-metacrista aproximadamente igual ou pouco menor que a pré-metacrista; protocrista como em M1; crista paraestilar como em M1; cíngulos como em M1; estilos como em M1; ectoestilo j podendo ser tão desenvolvido quanto l.
M3 - maior que os demais molares superiores; comprimento meso-distal da coroa pouco maior que M2; menor comprimento meso-dististilo, igual ou menor que a largura mesoestilo-protocone; bacia do cíngulo paraestilar aproximadamente igual à bacia do trígono, maior que M2 em área; bacia do cíngulo metaestilar entre o dobro e pouco menor em área que a bacia do cíngulo paraestilar; pós-paracrista entre igual e pouco menor que a pré-metacrista; protocrista e plagiocrista como em M1 e M2; crista paraestilar entre igual e dois terços da crista metaestilar; cíngulos como M1 e M2 quanto ao desenvolvimento; mesoestilo e dististilo como em M1 e M2; mesoestilo e dististilo reduzidos, mas sempre presentes; ectoestilos j e l bem desenvolvidos, j geralmente mais robustos, da altura do eocone; demais ectoestilos presentes ou ausentes, quando presentes geralmente mais evidentes que em M2.
M4 - o menor molar superior; comprimido no sentido antero-posterior do palato; assemelha-se a porção mesial dos demais molares superiores; comprimento meso-distal da coroa menor que nos demais molares superiores, pouco menor que a largura mesoestilo-eocone; largura mesestilo-protocone; maior que em M3 ; bacias paraestilar e metaestilar fundidas, a metaestilar bastante reduzida; bacia do trígono mais reduzida que nos demais molares, aproximadamente a metade; o eocone e o protocone mais desenvolvidos que o metacone; paracrista cinco ou seis vezes maior que a pós-metacrista; pós-metacrista e pré-metacrista muito reduzidas, a pré-metacrista metade ou um terço da pós-paracrista; protocrista geralmente mais curta que nos demais molares, plagiocrista igual ou mais curta; crista metaestilar reduzida; cíngulo bucal mais reduzido que nos demais molares superiores; mesoestilo e dististilo sempre presentes; ectoestilo j presente ou ausente, demais ectoestilos geralmente ausentes ou muito reduzidos.
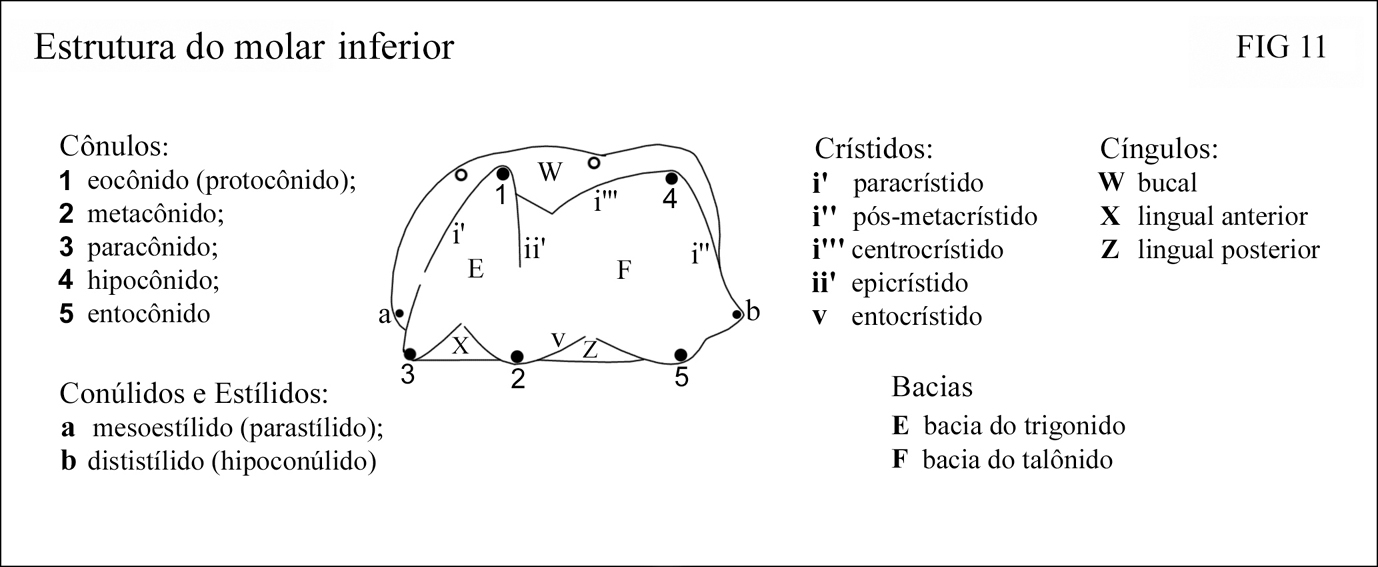
Molares inferiores m1, m2 e m3 semelhantes; m4 com trigonídeo como nos demais e talonídeo um pouco modificado; aproximadamente retangulares em vista oclusal e tridentados em vista bucal; comprimento aproximadamente o dobro da largura (fig. 9 e 11)
O rinário é a ponta do focinho. Esse órgão apresenta certa variação dentro do gênero. Nas espécies M. domestica, M. glirina, M. brevicaudata e M. amazonica, a borda dorsal posterior do rinário é reta; enquanto nas demais espécies projeta-se em ponta. Nas espécies M. brevicaudis e M. dimidiata essa ponta é bem evidente. A fig. 12 mostra rinários de algumas espécies em vista dorsal. A borda reta representa a condição plesiomórfica nos marsupiais, a projeção em ponta é derivada e é extrema em M. brevicaudis e M. dimidiata. No grupo M. emiliae essa borda é menos evidente que no grupo M. adusta.
Orelhas


As orelhas variam principalmente quanto ao tamanho relativo da hélix. Podemos classificá-las em quatro tipos:
(a) Grande, hélix bem expandida, em M. domestica.
(b) Média-grande, em M. glirina, M. brevicaudata e M. amazonica.
(c) Média-curta, em M. emiliae, M. scalops e M. americana.
(d) Curta, principalmente em M. brevicaudis,M. dimidiata e M. macae.
mas também em M. adusta, M. kunsi, M. osgoodi, M. obscura e M. iheringi.
A espécie M. obscura (fig. 13) apresenta uma pilosidade mais intensa nas orelhas, se comparada às demais espécies.

Patas
Anteriores (manus)
Anteriores (manus)
As almofadas plantares são maiores em M. domestica e relativamente reduzidas nas demais espécies, especialmente no grupo M. americana. Geralmente a almofada 1 é de tamanho médio, as almofadas 2, 3 e 4 são as menores, sub-iguais e a 5 é a maior; em alguns espécimes a almofada 1 pode estar dividida em duas menores, em Monodelphis amazonica é alongada.
O dedo V pode ser mais alongado e dobrável, com a ponta atingindo a segunda falange do dedo IV ou pode ser curto e reto com a ponta na altura da terceira falange do dedo IV. No grupo M. americana o dedo V encontra-se deslocado em direção a região carpal implantando-se aproximadamente na altura do dedo I (fig. 14).

Posteriores (pes)
As mesmas condições observadas nas patas anteriores também ocorrem nas posteriores. As almofadas são mais desenvolvidas em M. domestica e mais reduzidas no grupo M. americana. Em M. brevicaudata e M. amazonica, 1 e 2, são aproximadamente triangulares muito próximas uma da outra. Em M. obscura, 1 e 2 estão fundidos numa almofada única. O artelho V é um pouco mais deslocado para a região tarsal no grupo M. americana (fig. 15).
As unhas são mais curvas nos artelhos que nos dedos (fig. 2).
As mesmas condições observadas nas patas anteriores também ocorrem nas posteriores. As almofadas são mais desenvolvidas em M. domestica e mais reduzidas no grupo M. americana. Em M. brevicaudata e M. amazonica, 1 e 2, são aproximadamente triangulares muito próximas uma da outra. Em M. obscura, 1 e 2 estão fundidos numa almofada única. O artelho V é um pouco mais deslocado para a região tarsal no grupo M. americana (fig. 15).
As unhas são mais curvas nos artelhos que nos dedos (fig. 2).

Cauda
A variação da cauda está geralmente relacionada com o comprimento dos pelos das escamas (as tríades). Comparando os pelos laterais da porção mediana da cauda, M. glirina, M. americana, M. iheringi e grupo M. adusta possuem os pelos das tríades mais curtos, com um comprimento aproximadamente de duas escamas. O pelo central da tríade é geralmente mais grosso.
Em M. domestica a cauda pode estar um pouco mais coberta que nas espécies acima citadas, principalmente nas populações do nordeste brasileiro. Os pelos laterais das tríades geralmente são tão longos como o pelo central. O comprimento variando entre duas ou três escamas.
Nas espécies M. brevicaudis, M. dimidiata, Monodelphis amazonica e M. scalops os pelos da cauda variam de comprimento entre três ou quatro escamas.
A variação da cauda está geralmente relacionada com o comprimento dos pelos das escamas (as tríades). Comparando os pelos laterais da porção mediana da cauda, M. glirina, M. americana, M. iheringi e grupo M. adusta possuem os pelos das tríades mais curtos, com um comprimento aproximadamente de duas escamas. O pelo central da tríade é geralmente mais grosso.
Em M. domestica a cauda pode estar um pouco mais coberta que nas espécies acima citadas, principalmente nas populações do nordeste brasileiro. Os pelos laterais das tríades geralmente são tão longos como o pelo central. O comprimento variando entre duas ou três escamas.
Nas espécies M. brevicaudis, M. dimidiata, Monodelphis amazonica e M. scalops os pelos da cauda variam de comprimento entre três ou quatro escamas.
As espécies M. emiliae e Monodelphis obscura apresentam pelos relativamente longos para o gênero, aproximadamente cinco escamas de comprimento. Em Monodelphis obscura as escamas estão praticamente ausentes (fig. 16).
A cauda mais peculiar, por suas características, é a de M. brevicaudata. Nesta espécie os pelos da extremidade proximal dorsal, avançam pelo primeiro terço do comprimento da cauda, reduzindo gradativamente de tamanho. A parte ventral é coberta por pelos curtos (duas escamas de comprimento). Na porção mediana lateral os pelos centrais das tríades possuem um comprimento de aproximadamente quatro escamas (fig. 17). No entanto, apenas algumas populações de M. brevicaudata possuem esse padrão, as populações do oeste como M. b. palliolata e M. b. orinoci retêm o padrão geral do gênero.

Medidas corporais
De acordo com as medidas fornecidas pelos coletores, a espécie de maior tamanho é M. domestica e as menores M. iheringi e M. kunsi. O adulto de M. domestica podem chegar a 280 mm de comprimento total, M. iheringi atinge no máximo 160 mm, e M. kunsi 113 mm. A relação entre o comprimento cabeça-tronco e o comprimento da cauda é aproximadamente de dois para um na maioria das espécies. Ou seja, a cauda mede aproximadamente a metade do comprimento do corpo.
As espécies M. domestica, M. glirina e M. brevicaudata, possuem caudas relativamente mais compridas que as demais espécies. As espécies M. emiliae e M. scalops possuem as caudas mais curtas com relação as demais.
A relação entre a altura da orelha e o comprimento da cabeça mais o tronco, varia entre um quinto e um doze avos. Em M. domestica as orelhas são proporcionalmente maiores e em M. brevicaudis e M. dimidiata proporcionalmente menores.
De acordo com as medidas fornecidas pelos coletores, a espécie de maior tamanho é M. domestica e as menores M. iheringi e M. kunsi. O adulto de M. domestica podem chegar a 280 mm de comprimento total, M. iheringi atinge no máximo 160 mm, e M. kunsi 113 mm. A relação entre o comprimento cabeça-tronco e o comprimento da cauda é aproximadamente de dois para um na maioria das espécies. Ou seja, a cauda mede aproximadamente a metade do comprimento do corpo.
As espécies M. domestica, M. glirina e M. brevicaudata, possuem caudas relativamente mais compridas que as demais espécies. As espécies M. emiliae e M. scalops possuem as caudas mais curtas com relação as demais.
A relação entre a altura da orelha e o comprimento da cabeça mais o tronco, varia entre um quinto e um doze avos. Em M. domestica as orelhas são proporcionalmente maiores e em M. brevicaudis e M. dimidiata proporcionalmente menores.
Para continuar clique neste LINK
Nenhum comentário:
Postar um comentário
Observação: somente um membro deste blog pode postar um comentário.